平成23年度厚生労働科学研究費補助金
障害者対策総合研究推進事業(感覚器障害分野)報告書
ラット音響外傷モデルを用いた難聴発現の解析
吉田 正
市立宇和島病院
1.はじめに
老人性難聴は高齢化社会が進んだ本邦において益々増加する疾患と考えられる。高齢者にとって難聴はコミュニケーションを阻害し、QOLを低下させる。超磁歪素子を用いた埋め込み型骨導補聴器は、700万人と推定される、これら老人性感音難聴患者にも適応拡大が可能なデバイスである。このデバイスの臨床応用に向けて感音難聴発現のメカニズムを解析することは非常に重要である。
近年、強大音曝露による易受傷性と加齢に伴う聴覚閾値の変化(老人性難聴)との関連性が報告されている。そこで、老人性難聴の進行に関するメカニズムを解明するために音響外傷モデルを用いて実験を行った。強大音響負荷による難聴発現のメカニズムには、内耳に対する機械的障害と代謝的障害の2種類の機序が考えられているが、その詳細な機序は未だ不明である。今回、2つの異なる系統のラットに対して音響外傷モデルを作成し、それぞれの蝸牛におけるABRによる聴力評価およびマイクロアレイを用いた遺伝子発現の解析を行った。
2.方法
8~10週齢の雌のDA系ラットとPVG系ラットを本実験に用いた。
4kHzを中心とした広帯域ノイズ(帯域幅3.2kHz)に110dBの音圧レベルで4時間曝露した。騒音曝露前、曝露24時間後および72時間後に、3.5、7、14および28kHzのABR閾値を測定した。
各時点において、コントロールのラット3匹および曝露後のラット9匹(DA系ラット4匹およびPGV系ラット5匹)の蝸牛をマイクロアレイ解析に用いた。各時点でのコントロール群と実験群およびDA系ラットとPVG系ラット間の比較は、標準化後にt-testを用いて行った。異なるカットオフ閾値毎にfalse discovery rate(FDR)を算出し、FDR<0.05のプローブセットは有意に発現量が変化しているとみなした。これらの遺伝子については、Database for Annotation,Visualization and Integrated Discovery(DAVID)を用いて更に解析を行った。
3.結果および考察
曝露前の3.5、7、14および28kHzでの平均ABR閾値は、DA系ラットでそれぞれ23.2±3.0dB、14.1±3.2dB、10.9±2.6dBおよび18.8±4.2dBであり、PVG系ラットでそれぞれ23.4±2.4dB、15.6±3.1dB、12.5±3.2dBおよび20.9±3.3dBであった。いずれの周波数においても2つの系統間で有意差は見られなかった。騒音曝露後には両系統ともABR閾値は上昇した(図1)。曝露24時間後の閾値上昇変化は、すべての周波数においてDA系ラットに比べてPVG系ラットで有意に大きかった(図1A)。曝露72時間後では、両系統おいて閾値上昇の抑制つまりABR閾値の回復は限定的で、すべての周波数において閾値上昇が持続していた(図1B)。PGV系ラットにおいて、曝露72時間後のABR閾値の回復は有意に乏しかった。
マイクロアレイ解析においてバッチ効果の調整を行った後、すべてのサンプルはDA系ラットとPVG系ラット間で明確な分離が見られた。表1は標準化後、有意に発現量が変化したプローブセットの数を示している。曝露72時間後のDA系ラットとPGV系ラット間では有意差を認めたが、他のすべての時点および系統内では有意差を認めなかった。有意差を認めた曝露72時間後におけるDA系対PVG系の1439個のプローブセットについては、DAVIDシステムを用いて更に解析を行った。表2は機能予測チャートを示しているが、その結果、これらの発現量の変化した遺伝子は、炎症反応に強く関連していた。
データの分析に関しては更に詳細な検討が必要であるが、これまでの結果から音響外傷における内耳障害には炎症反応に関係のある遺伝子が強く関与していることが示唆された。よって音響外傷と関連性がある老人性難聴においても、その難聴の進行に炎症に関する遺伝子が関与していることが推測される。今後、加齢性難聴モデルでも同様の研究を行い、難聴発現のメカニズムを解析することが望まれる。これにより難聴の進行を防御する手段の手掛かりを得ることができ、超磁歪素子を用いた埋め込み型骨導補聴器の適応拡大が期待できると考える。
図1 騒音曝露の24時間および72時間後におけるABR閾値変化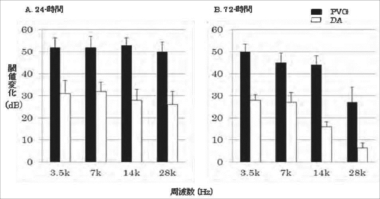 (図1の内容)
(図1の内容)
表1 有意に発現量が変化したプローブ数(FDR<0.05)
| Time and strain | Number of probe sets | Annotated genes* |
|---|---|---|
| DA24hrs(vs cont) | 0 | 0 |
| DA24hrs(vs cont) | 1 | 1 |
| PVG24hrs(vs cont) | 0 | 0 |
| PVG72hrs(vs cont) | 0 | 0 |
| Control(DA/PVG) | 0 | 0 |
| 24hrs(DA/PVG) | 0 | 0 |
| 2hrs(DA/PVG) | 1439 | 975 |
表2 72時間時点におけるDA系対PVG系の機能予測チャート
975遺伝子(274遺伝子で発現上昇し、596遺伝子で発現減少)
| category | Term | count | PValue | Bonferroni | Benjamini |
|---|---|---|---|---|---|
| GOTOERM_BP_FAT | Immuneresponse | 61 | 4.95E-15 | 1.40E11 | 1.40E11 |
| GOTOERM_BP_FAT | Antigen processing and presentation | 20 | 7.64e-09 | 2.14E-05 | 1.07E-05 |
| GOTOERM_BP_FAT | Antigen processing and presentation of peptide antigen | 14 | 1.38E-08 | 3.88E-05 | 1.29E-05 |
| GOTOERM_BP_FAT | Antigen processing and presentation of peptide antigen via MHC class1 | 11 | 3.45E-08 | 9.69E-05 | 2.42E-05 |
| SP_PIR_KEYWORDS | MHC1 | 9 | 8.23E-07 | 3.00E-04 | 3.00E-04 |
| GOTOERM_BP_FAT | Immune effectorprocess | 21 | 9.32E-07 | 0.00261 | 5.23E-04 |
| SP_RIP_KYEWORDS | Immune response | 19 | 1.55E-06 | 5.64E-04 | 2.82E-04 |
| GOTOERM_BP_FAT | Defense response | 42 | 2.39E-06 | 0.00668 | 0.001116 |
| INTERPRO | MHC class I,alpha chain,C-terminal | 8 | 3.02E-06 | 0.003038 | 0.003038 |
| GOTERM_CC_FAT | MHC class Iprotein comples | 11 | 3.20E-05 | 0.013393 | 0.013393 |
| GOTOERM_BP_FAT | Prositive regulation of immune system process | 27 | 3.99E-05 | 0.105814 | 0.01585 |
| GOTOERM_BP_FAT | Response to virus | 11 | 4.58E-05 | 0.120635 | 0.015941 |
| KEGG_PATHWAY | Antigen processing and presentation | 16 | 4.60E-05 | 0.00747 | 0.00747 |
| INTERPRO | MHC class I,alpha chain,alpha1 and alpha2 | 10 | 6.90E-05 | 0.067057 | 0.03411 |
| INTERPRO | MHC class I-like antigen recognition | 10 | 1.71E-04 | 0.157959 | 0.055698 |